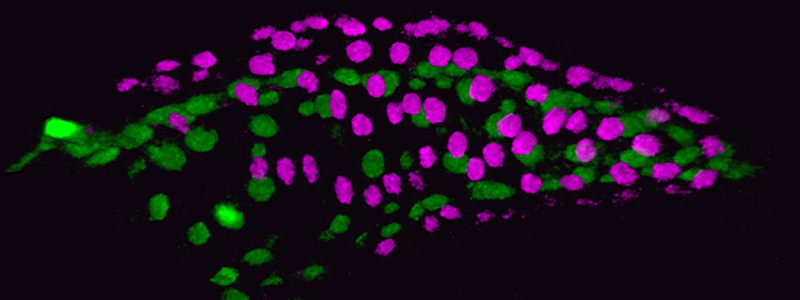
Developing Zebrafish Heart

A young freshwater turtle

Carnivorous flowering plant

Wildflower diversity in a restored open oak woodland

Diversity of wild mushrooms of north Mississippi

Savannah meadowbeauty (Rhexia alifanus)

Young shark being studied in the Gulf of Mexico
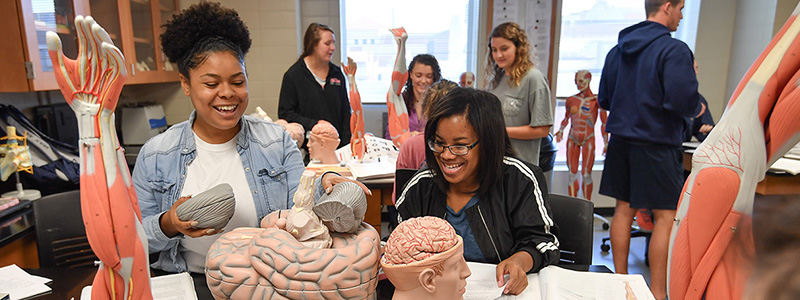
Anatomy & Physiology laboratory in Shoemaker Hall

Zebra finch neurobiology research

Tarantula photographed in Arizona

Visualization of Reactive Oxygen Species in Guard Cells
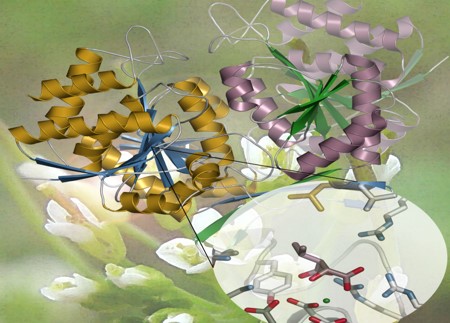
The first crystallized plant isopropylmalate dehydrogenase







